Research Article
Volume 7 Issue 1 - 2025
Biophysical characterization of nesting sites of olive ridley turtle (Lepidochelys olivacea, Eschsholtz, 1829) in the «Manyange na Elombo-Campo» National Marine Park, South Cameroon (West-Africa)
1Department of Forestry, Laboratory of Ichthyology and Applied Hydrobiology, University of Dschang, Cameroon
2Department of Oceanography, Institute of Fisheries and Aquatic Sciences, University of Douala, Cameroon
3Local Community Research and Development Association Tube Awù, South Cameroon Region, Cameroon
4Sea Turtle Research Centre, Mas du Ringué, 46260 Beauregard, France
2Department of Oceanography, Institute of Fisheries and Aquatic Sciences, University of Douala, Cameroon
3Local Community Research and Development Association Tube Awù, South Cameroon Region, Cameroon
4Sea Turtle Research Centre, Mas du Ringué, 46260 Beauregard, France
*Corresponding Author: Nana Towa Algrient, Department of Forestry, Laboratory of Ichthyology and Applied Hydrobiology, University of Dschang, Cameroon. Email Id: algrient@yahoo.fr
Received: January 24, 2025; Published: February 03, 2025
Abstract
In Cameroon, the «Mayange na Elombo-Campo National Marine Park» has been identified as one of the regular and stable nesting and feeding sites for marine turtles. However, the latter is subject to natural and anthropogenic threats which could later lead to their alteration as well as to the disappearance of the species as Lepidochelys olivacea. The objective of the present study is to contribute to the conservation of this specie of marine turtle through a better knowledge of the biophysical characteristics of its nesting sites in the «Manyange na Elombo-Campo National Marine Park». To achieve this, survey for direct observations, consultation of documents and topographic survey using a GPS were carried out. The study revealed that the back beaches where marine turtles nest are inhabited by 9 plant species grouped into 6 families. The animal species that frequent these sites belong to 4 classes, namely crabs, mammals, birds and reptiles. The substrate of the sites is homogeneous and sandy in nature and the grain size reveals three types of sand; fine, medium and coarse whose sediment sizes are respectively >60 to < 465 µm, >465 to < 2000 µm, and > 2000 to < 4750 µm. The morphology (slope, highest and lowest altitude point, width and accessibility) of the sites differs depending on the nesting sites. Particle size shows a significantly high correlation with hatching (r²=0.95) and emergence (r²=0.97) and therefore significantly influences incubation success.
Keywords: Biophysical characterization; Marine turtle; Nesting site; National Marine Park
Introduction
Marine turtles are reptiles that spend the majority (99%) of their lives at sea and are important consumers within trophic chains Bjorndal & Jackson, 2003. This role as consumers is important for the regulation of their prey Bjorndal et al, 2000, and as bio indicators, they provide information on different compartments of marine ecosystems. Thus, they can be qualified as umbrella species, meaning that their protection could promote the protection of a large number of other species sharing the same habitats Bourges, 2000. Just like birds and certain marine mammals, they ensure their reproduction on the continent (nesting sites). « A nesting site for marine turtles is any coastal land area where at least one female of any species has laid eggs in historical times. » Fretey & Girondot, 1996.
This terrestrial nesting habitat includes the three littoral zones of the emerged part, more or less long, of the beaches: infralittoral zone, intertidal or mediolittoral zone (foreshore), supralittoral zone. The locomotion of an adult female turtle (mainly Eretmochelys imbricata, but also sometimes Lepidochelys olivacea) can lead it to overshoot its usual floor, either voluntarily or by accident (artificial lights causing disorientation). Females D. coriacea frequent terrestrial habitats accessible via deep water and strong waves, devoid of rocky mass, beach-rocks or any other abrasive obstacle that could cause injury Fretey & Triplet, 2021.
Nesting sites are made up of two main areas: the beach itself and the backshore. The beach is divided into two sub-units: the beach of sand or other types of substrate, and an upper part of the beach more or less covered with low, creeping herbaceous vegetation. These beaches are subject to a number of physical threats, mainly natural erosion or erosion caused by port infrastructures, leading to a narrowing of the sandy strip Fuentes et al. These spaces undergo sediment erosion dynamics and strong anthropogenic pressure on the terrestrial domain, leaving little space for natural sedimentary processes Defeo et al., important for the nesting of marine turtles. Environmental parameters linked to the quality and type of substrate such as temperature, topography (slope of the beach, presence of natural barrier), particle size of the substrate, geographical location of the nest (exposure to swell, presence of vegetation), are determinants in the ultimate choice of the nesting site and the egg-laying decision by gravid females once they arrive at the nesting site Wood & Bjorndal, 2000.
In Cameroon, the “Manyange na Elombo Campo National Marine Park”, on the northern border with Equatorial Guinea, has been identified as an important nesting and feeding site for marine turtles. The park's nesting sites are home to two species of marine turtle: the Olive ridley, the Leatherback and, more sporadically, the Green Turtle. The coastal areas are frequented by juvenile Green Turtles and Hawksbill Turtles.
However, these sites are subject to natural and anthropogenic threats which could later lead to their alteration as well as the disappearance of these species; these include, among other things, pollution by tree species lying on the site; poaching; predation by animal species that frequent nesting beaches; the shrinkage of the sandy strip IUCN, 2012; flooding of nesting sites due to high tides; coastal erosion.
The present study aimed to contribute to the conservation of marine turtles through a better knowledge of the biophysical characteristics of nesting sites in the “Manyange na Elombo-Campo Marine National Park”. Specifically, these are:
- Determine the composition of animal species and woody species present on marine turtle nesting sites
- Identify the nature, determine the particle size of the substrate and the topo-morphology of marine turtle nesting sites
- Evaluate the influence of particle size on the success of incubation of sea turtle eggs.
Material and Methods
Presentation of the Manyange na Elombo Campo Marine National Park
Created in 2021 by decree N°. 2021/4804/PM of 9 July 2021, the «Manyange na Elombo-Campo National Marine Park» is located in Cameroon's South Region, Ocean Department and Campo District. This marine park is adjacent to the maritime border with Equatorial Guinea, separated from the Rio Campo reserve by the Ntem River. It is Cameroon's first maritime and coastal protected area. It occupies the Kribi-Campo coastal strip and covers an area of 110,300 ha. It extends 42.364 km (26.5 miles) offshore, in Cameroonian territorial waters, towards the Equatorial Guinean Island of Bioko, and stops 300 meters from the coastal line of the highest tides. The park has a buffer zone of three thousand four hundred hectares (3,400 ha) reserved for community activities not harmful to biodiversity, with a maximum depth of 350 meters, bordering the marine national park on an immediate and intact coastal zone. This zone is delimited on the mainland, depending on the case, by a strip 50 or 25 meters after the highest tide line.
Created in 2021 by decree N°. 2021/4804/PM of 9 July 2021, the «Manyange na Elombo-Campo National Marine Park» is located in Cameroon's South Region, Ocean Department and Campo District. This marine park is adjacent to the maritime border with Equatorial Guinea, separated from the Rio Campo reserve by the Ntem River. It is Cameroon's first maritime and coastal protected area. It occupies the Kribi-Campo coastal strip and covers an area of 110,300 ha. It extends 42.364 km (26.5 miles) offshore, in Cameroonian territorial waters, towards the Equatorial Guinean Island of Bioko, and stops 300 meters from the coastal line of the highest tides. The park has a buffer zone of three thousand four hundred hectares (3,400 ha) reserved for community activities not harmful to biodiversity, with a maximum depth of 350 meters, bordering the marine national park on an immediate and intact coastal zone. This zone is delimited on the mainland, depending on the case, by a strip 50 or 25 meters after the highest tide line.
This zone extends over a width of 300 m into the sea and also covers the southern confluence of the Tyenjè watercourse with the Atlantic Ocean, the Cameroonian part of the mouth of the Ntem, Lake Pitché and the mangrove there adjoining (Figure 1).
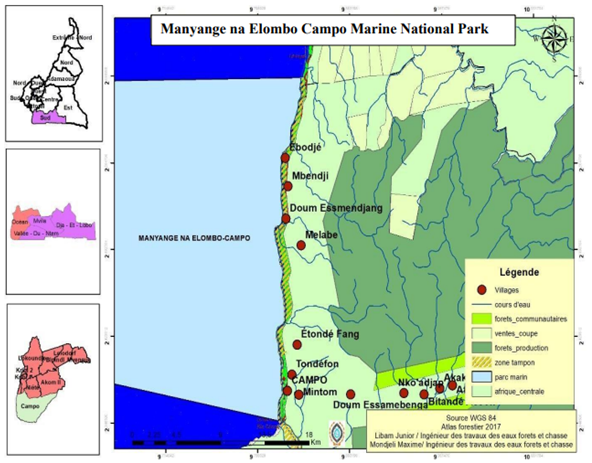
Figure 1: Study area.
Presentation of the Olive Ridley nesting sites
To the south of the National Marine Park, according to the local research and development association “Tubè Awù”, the nesting sites are named after outlying villages from Ebodjé to Campo via Bouandjo. The sites used in this study are Ebodjé, Bouandjo and Mbondo (Figure 2).
To the south of the National Marine Park, according to the local research and development association “Tubè Awù”, the nesting sites are named after outlying villages from Ebodjé to Campo via Bouandjo. The sites used in this study are Ebodjé, Bouandjo and Mbondo (Figure 2).
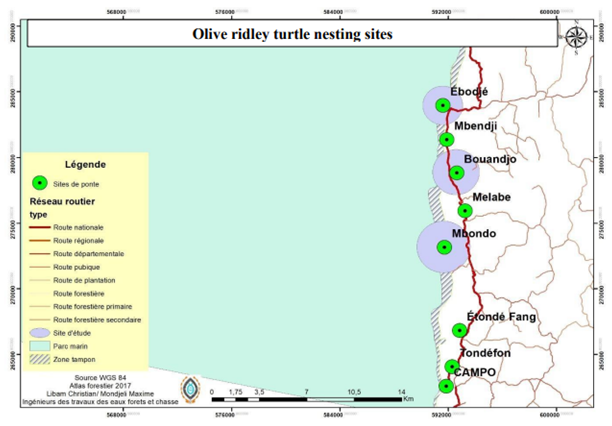
Figure 2: Olive Ridley turtle nesting sites in south of Ebodjé.
Collection of data
Determination of animal and woody species of marine turtle nesting sites
A structured interview was carried out with fishermen randomly selected from the different villages of the study sites. Ebodje (35 fishermen), Bouandjo (20 fishermen) and Mbondo (15 fishermen) for a total of 70 fishermen. In addition to surveys, direct observations on the beaches were made by patrols (during the day and at night depending on low tides). Animal species were identified using the French/Iyasa zoological lexicon for vertebrates by Fretey, 2003.
A structured interview was carried out with fishermen randomly selected from the different villages of the study sites. Ebodje (35 fishermen), Bouandjo (20 fishermen) and Mbondo (15 fishermen) for a total of 70 fishermen. In addition to surveys, direct observations on the beaches were made by patrols (during the day and at night depending on low tides). Animal species were identified using the French/Iyasa zoological lexicon for vertebrates by Fretey, 2003.
The quantitative inventory of Ocypode cursor crabs and other crab species frequenting the spawning sites was carried out at night in 50 x 10 m plots, covering an area of 500 m². The technique used was the artisanal fishing technique of the Iyasa community called «Mangongo». This technique consisted of escaping containers which served as traps into the sand. This technique consisted of placing containers in the sand to act as traps. To do this, it was enough to take per plot 3 containers of 15 litters with openings of diameter 30 cm and height 35 cm, then dig three holes in the sand equal to the height of the containers and equidistant by 15 m. Then, insert the containers into the holes, cover the opening of the container with green and light leaves. Bait, consisting of cassava flour or grated coconut above and around the containers. The species of crabs which live behind the beach and regularly frequent the nesting beaches were captured. All these crab species were identified using the publication for West African Brachyuran Crabs Manning & Holthuis, 1981 and the identification guide for Lagoon Crabs in South Benin, West Africa Dessouassi et al. 2018. The calculated parameter is the frequency of observation of each species on the nesting site.
The botanical inventory method consisted of inventorying tree and shrub species according to Angoni, 2005. Trees with a diameter at breast height (DBH) of at least 10 cm were recorded. The botanical inventory system was based on the establishment of three rectangular plots of 50x10 m, i.e. an area of 500 m² equidistant by one km per transect, symmetrical to the plots used for the inventory of crab species in relation to the vegetation line. The species were identified using the Picture and Plantnet applications and then revised by a botanist from the Mbalmayo National School of Water and Forests. The calculated parameter is the specific density of each species.
Identification of the nature of the substrate and determination of the grain size and topo-morphology of marine turtle nesting sites
Samples of between 500g and 1000g of sand were taken from the plots (used for the crab inventory), and preserved in A4 format plastics. Three samples were taken per transect, for a total of nine samples. These samples were analyzed at the soil science laboratory of the Faculty of Agronomy and Agricultural Sciences of the University of Dschang using the sieving method. The sieves used had respective mesh sizes of 60, 400, 2000, 5000 micrometers (0.06; 0.4; 2; 5 mm) and the visual identification key for sand was used to classify the categories of sand obtained.
Samples of between 500g and 1000g of sand were taken from the plots (used for the crab inventory), and preserved in A4 format plastics. Three samples were taken per transect, for a total of nine samples. These samples were analyzed at the soil science laboratory of the Faculty of Agronomy and Agricultural Sciences of the University of Dschang using the sieving method. The sieves used had respective mesh sizes of 60, 400, 2000, 5000 micrometers (0.06; 0.4; 2; 5 mm) and the visual identification key for sand was used to classify the categories of sand obtained.
The morphology of the nesting sites (the slope, the highest point and the lowest point) was measured by a topographic survey using the Garmin etrex 10 GPS device. The data was recorded in three dimensions: latitude, longitude and altitude and processed using Global maper and Google Heart software to generate the morphology of the sites.
Evaluation of the influence of particle size on the incubation success of sea turtle eggs
The nests obtained in this study belong to those of the olive ridley turtle. The nests were identified using footprints left on the sand by the species, and by direct observation of the species laying eggs (Figure 3).
The nests obtained in this study belong to those of the olive ridley turtle. The nests were identified using footprints left on the sand by the species, and by direct observation of the species laying eggs (Figure 3).

Figure 3: Footprint of Olive Ridley (a) and nesting activity of Olive Ridley (b).
The 12 nests of Olive Ridley used in this study were nests transplanted in situ (in the same natural environment). Incubation success was determined by the nest profile methodology according to Bell et al.; Maite & Agambouet, 2008. The nests were excavated two weeks after the probable hatching period. Once excavated, the contents of the nests were inspected, the number of eggs that failed to hatch was counted and the number of hatchlings that failed to emerge was also counted.
Data analysis
The data collected in this study were encoded and processed using the Excel spreadsheet. The Google Maper and Google Earth software were used to highlight the morphology of the sites and maps were produced by the Arcgis mapping software. The statistical test used to show the correlation between particle size and incubation success was that of Pearson carried out using SPSS 20.0 software.
The data collected in this study were encoded and processed using the Excel spreadsheet. The Google Maper and Google Earth software were used to highlight the morphology of the sites and maps were produced by the Arcgis mapping software. The statistical test used to show the correlation between particle size and incubation success was that of Pearson carried out using SPSS 20.0 software.
Results
Composition of animal species and woody species encountered on olive ridley turtle nesting sites
It appears from this table that, the spawning sites are composed of six species of crabs grouped into two families: Ocypodideae and Grapsydeae. Mammals are made up of seven species, reptiles made up of four species, and birds made up of six species.
It appears from this table that, the spawning sites are composed of six species of crabs grouped into two families: Ocypodideae and Grapsydeae. Mammals are made up of seven species, reptiles made up of four species, and birds made up of six species.
| Crabs | |||
| Family | Scientific name | Common name | Vernacular Name |
| Ocypodideae | Ocypode cursor Linnaeus, 1758 | Livid ghost Crab | Koko a monanga, mêmè |
| Ocypode gaudichaudii Eydou, 1835 | Galapagos ghost crab | Koko a ngoyè, popi | |
| Ocypode cordimanus Latreille, 1818 | Brown Earth Crab | Jombo | |
| Uca neocultrimana Bott, 1973 | Common fidler crab | Vikopida | |
| Grapsideae | Geograpsus grayi H. Milne Edwards, 1853 | Rough ground crab | Jàlà |
| Pachygrapsus planifrons De Man 1888 | Common runner crab | Dangadaya | |
| Other classes | |||
| Mammals | Reptiles | Birds | |
| Dog | Monitor lizard | Crows | |
| Civet | Wooden snake | Oxpecker | |
| Pangolin | Agama lizard | Knight, Little puppet | |
| Hedgehog | Viper | Sparrow, hawk | |
| Palm rat | Eagle | ||
| Striped cat | |||
| Monkey | |||
Table 1: List of animal species on nesting sites obtained from surveys and documents.
| Crustaceans | Mammals | Birds | Reptiles |
| Crabs | Dog | Knight | Agama lizard |
| Civet | Crow | Monitor lizard | |
| Pangolin Monkey | Sparrow Hawk | ||
| Oxpecker |
Table 2: List of animal species observed directly on nesting sites.
Composition and relative density of woody species
The results obtained in the field made it possible to determine the composition of the woody species of the nesting sites and at the same time to highlight the different relative densities of these species per nesting site. In total, nine woody species were identified and divided into six families. Thus, the Clusiaceae family consists of a single species of Calophyllum inophyllum and offers the highest average relative density of 58.35%. Then comes the Arecaceae family with an average relative density of 13.63%, composed of Cocos nucifera, Elaeis guineensis Phoenix. sp; The Moraceae family with an average relative density of 8.83% is composed of species of the Ficus genus. The Combretaceae family is composed of a single woody species Terminalia catapa with an average relative density of 6.61%. The families Sapotaceae (11.07%) and Urticaceae (1.52%) are composed respectively of the species Manilkara abovata and Musanga cecropioides.
The results obtained in the field made it possible to determine the composition of the woody species of the nesting sites and at the same time to highlight the different relative densities of these species per nesting site. In total, nine woody species were identified and divided into six families. Thus, the Clusiaceae family consists of a single species of Calophyllum inophyllum and offers the highest average relative density of 58.35%. Then comes the Arecaceae family with an average relative density of 13.63%, composed of Cocos nucifera, Elaeis guineensis Phoenix. sp; The Moraceae family with an average relative density of 8.83% is composed of species of the Ficus genus. The Combretaceae family is composed of a single woody species Terminalia catapa with an average relative density of 6.61%. The families Sapotaceae (11.07%) and Urticaceae (1.52%) are composed respectively of the species Manilkara abovata and Musanga cecropioides.
The species Calophillum inophillum, Cocos nucifera, Terminalia catapa are present on the three sites but are more abundant on the Bouandjo site with relative densities respectively of 74.13%; 11.43 and 7.62%. The species Elaeis guineensis (9.3%) is only present on the Ebodje site; the species Phoenix. sp (4.19%) is only present on the Bouandjo site; the species Ficus benjamina (4.74%) is only present on the Mbondo site, on the other hand the species Ficus saussureana is present on the Ebodje and Mbondo sites but, more abundant on the Ebodje site and offers a relative density of 17.15%, the species Manilkara abovata is present on the three sites but is more abundant on the Ebodje site and offers a relative density of 26.43%; however, the species Musanga cecropioides (4.55%) is only present on the Mbondo site.
The shrubs on the plots are essentially made up of the species Calophyllum inophyllum, hence its high relative density; moreover, the waterfront is mainly made up of this species, followed by the species Manilkara abovata, Cocos nucifera, Terminalia catapa and Ficus saussureana.
| Relative density (%) | |||||
| Families | Woody species | Sites | ARD | ||
| Ebodje | Bouandjo | Mbondo | |||
| Arecaceae | Cocos nucifera | 11.32 | 11.43 | 4.65 | 9.13 |
| Elaeis guineensis | 9.3 | 0 | 0 | 3.10 | |
| Phoenix sp. | 0 | 4.19 | 0 | 1.40 | |
| Total | 20.62 | 15.62 | 4.65 | 13.63 | |
| Clusiaceae | Calophyllum inophyllum | 31.17 | 74.13 | 69.74 | 58.35 |
| Total | 31.17 | 74.13 | 69.74 | 58.35 | |
| Combrataceae | Terminalia catapa | 4.63 | 7.62 | 7.58 | 6.61 |
| Total | 4.63 | 7.62 | 7.58 | 6.61 | |
| Moraceae | Ficus benjamina | 0 | 0 | 4.74 | 1.58 |
| Ficus saussureana | 17.15 | 0 | 4.6 | 7.25 | |
| Total | 17.15 | 0 | 9.34 | 8.83 | |
| Sapotaceae | Manilkara abovata | 26.43 | 2.64 | 4.13 | 11.07 |
| Total | 26.43 | 2.64 | 4.13 | 11.07 | |
| Urticaceae | Musanga cecropioides | 0 | 0 | 4.55 | 1.52 |
| Total | 0 | 0 | 4.55 | 1.52 | |
ARD = Average Relative density
Table 3: Composition and relative density of woody species according to egg-laying sites.
Table 3: Composition and relative density of woody species according to egg-laying sites.
Nature and particle size of turtle nesting substrate
After sieving the substrate samples from the three sites, it appears that the size of the sediments is not less than 60 µm, which allows us to conclude that the sites are composed of a homogeneous substrate, of a sandy nature (Table 3).
After sieving the substrate samples from the three sites, it appears that the size of the sediments is not less than 60 µm, which allows us to conclude that the sites are composed of a homogeneous substrate, of a sandy nature (Table 3).
| Type of sand | Particle size class (µm) |
| Fine sand | 60-465 |
| Medium sand | 465-2000 |
| Coarse sand | 2000-4750 |
Table 4: Nature and particle size of turtle nesting substrate.
Overall, it appears from Table 3 that the substrate ranges from fine to coarse sand, belonging to the respective particle size classes >60 and465 and2000 and
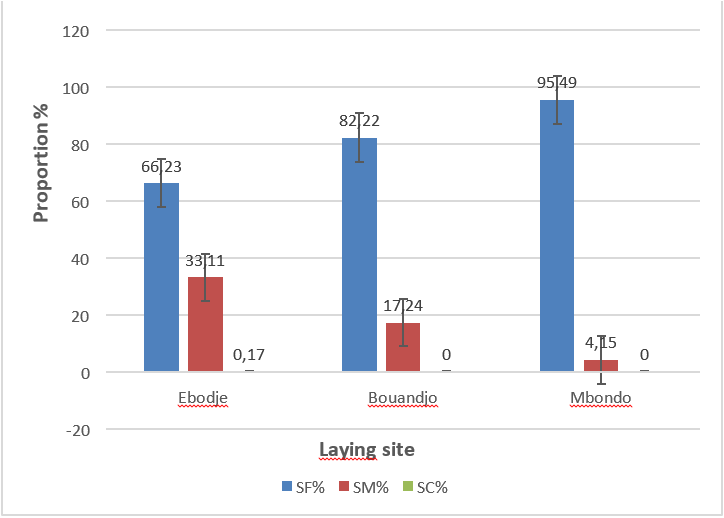

Proportion of particle size classes according to laying sites Figure 4 below shows the behavior of the different categories of sand on the spawning sites. It appears that fine sand is dominant on all the nesting sites of the marine park, because its proportion varies from 66.23% to 95.49%. On an equal site, fine sand is mainly represented at Mbondo with a proportion of 95.49% followed by the Buandjo site 82.22%.
Legend: SF= fine sand; SM= medium sand; SC= coarse sand
Figure 4: Behaviour of different categories of sand on nesting sites.
Figure 4: Behaviour of different categories of sand on nesting sites.
Topo-morphology of nesting sites
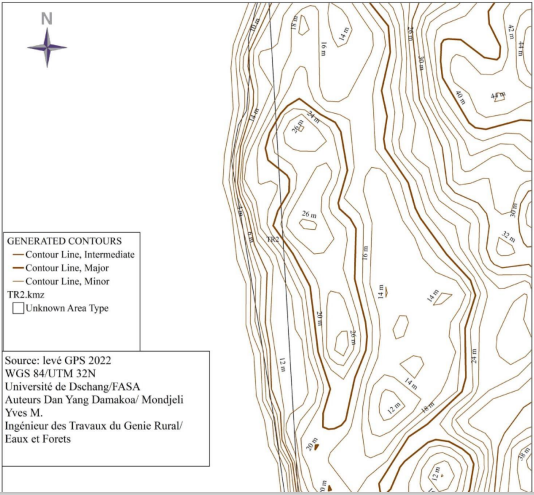
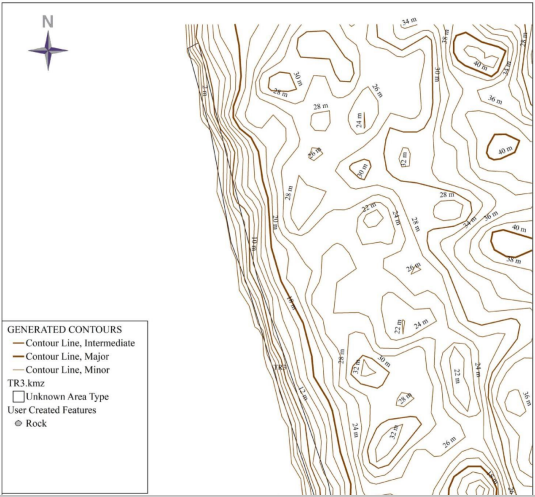
The topo-morphology of the nesting sites is illustrated in Figure below, the topo-morphological maps which highlight the quality of the slope, give an idea of the accessibility of the environment, the width and give the highest and lowest altitude points of the Ebodje nesting sites, Bouandjo and Mbondo. These maps present three different contour lines, major, intermediate and lower or minor.
The topo-morphology of the nesting sites is illustrated in Figure below, the topo-morphological maps which highlight the quality of the slope, give an idea of the accessibility of the environment, the width and give the highest and lowest altitude points of the Ebodje nesting sites, Bouandjo and Mbondo. These maps present three different contour lines, major, intermediate and lower or minor.
The Ebodje site has rocky areas, so access is virtually impossible in these parts of the site for marine turtles, the contour lines are very tight and equidistant throughout the site, which allows us to say that the site has a steep slope and regular (0.11-0.27). The width of the beach of this site ranges from 15 to 40m, the highest altitude point is 32m and the lowest is 2m and therefore the rise of sea turtles is difficult in this site. The Bouandjo site on the other hand does not have rocky areas, moreover the contour lines are not tight and are equidistant to the South of the site and tighten slightly towards the bottom of the North side which allows us to say that the slope (0, 10-0.16) is weak, regular and convex. The width of the beaches at this site ranges from 21 to 50m, the highest altitude point is 14m and the lowest is 2m. The sea turtle can easily access this site.
The Mbondo site has a rocky area to the south of the site, inaccessible for sea turtles. The contour lines are loose and equidistant throughout the site, which allows us to say that the site has a slight and regular slope (0.8-0.12). The width of the beaches on this site varies between 13 and 35 m, the highest altitude point is 10 m and the lowest is 2 m. the sea turtle easily accesses this site. All these morphological parameters are summarized in Table 4 below.
| Sites | Slope value (%) | HP (m) | LP (m) | Width (m) | Accessibility |
| Ebodje | 0.11-0.27 | 32 | 2 | 15-40 | Difficult to access |
| Bouandjo | 0.10-0.16 | 14 | 2 | 21-50 | Easily accessible |
| Mbondo | 0.8-0.12 | 10 | 2 | 13-35 | Easily accessible |
Legend: HP=Highest point, LP=Lowest point. Table 5: Morphological parameters of marine turtle nesting sites.
Morphology of the Ebodjé site
Morphology of the Bouandjo site
Morphology of the Mbondo site
Figure 5: Morphological maps of nesting sites.
Influence of particle size on nest incubation success
Characterization of nests
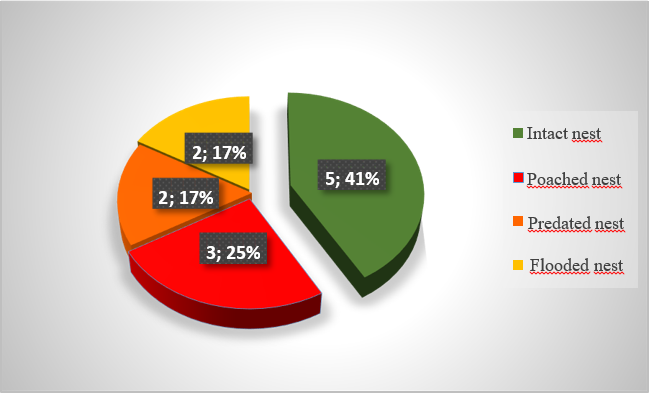
Of the twelve nests marked during this study, 5 nests (41%) produced at least one hatchling living outside the nest, and the remaining 7 (59%) had neither hatching nor emergence of twisters (nests destroyed). The causes of non-emergence are predation (17%), flooding by high tides (17%) and poaching (25%). Poaching is considered to be the greatest threat to marine turtle reproduction in this region (Figure 6).
Characterization of nests
Of the twelve nests marked during this study, 5 nests (41%) produced at least one hatchling living outside the nest, and the remaining 7 (59%) had neither hatching nor emergence of twisters (nests destroyed). The causes of non-emergence are predation (17%), flooding by high tides (17%) and poaching (25%). Poaching is considered to be the greatest threat to marine turtle reproduction in this region (Figure 6).
Figure 6: State of the nests obtained.
Analysis of the contents of nests found intact
Successful nests, found intact, were excavated and the contents analyzed. The following table 5 presents the classification of nest contents. It appears that the unhatched eggs representing 11.11% are classified as without apparent development because there is no visible embryo or bloodstains. 6.63% of unhatched eggs have holes in the membrane and are classified as predated. 4.09% are term fully formed embryos that have failed to emerge from the egg due to unknown causes. 6.04% of eggs are classified as “indeterminate” their contents being impossible to discern due to rotting.
Successful nests, found intact, were excavated and the contents analyzed. The following table 5 presents the classification of nest contents. It appears that the unhatched eggs representing 11.11% are classified as without apparent development because there is no visible embryo or bloodstains. 6.63% of unhatched eggs have holes in the membrane and are classified as predated. 4.09% are term fully formed embryos that have failed to emerge from the egg due to unknown causes. 6.04% of eggs are classified as “indeterminate” their contents being impossible to discern due to rotting.
| Classification of the contents of the 5 nests analyzed (N= 513) | ||
| Restrained new borns | 68 (13.26%) | Hatched eggs |
| Emergence | 302 (58.87%) | (N= 370) |
| Unhatched full-term embryos | 21 (4.09%) | Eggs no |
| Without apparent development | 57 (11.11%) | hatched (N=143) |
| Undetermined | 31 (6.04%) | |
| Unhatched eggs and predates | 34 (6.63%) | |
N= number of eggs
Table 6: Classification of contents of intact nests.
Table 6: Classification of contents of intact nests.
Success in incubating turtle eggs
When we consider only the unattacked nests, it appears overall that hatching success was on average 70.45% and emergence success 56.40% (table 6)
When we consider only the unattacked nests, it appears overall that hatching success was on average 70.45% and emergence success 56.40% (table 6)
| Code of Nests | Incubation of Nests | |
| Outbreak Achievement | Emergence Success | |
| N°1 | 81.25 | 76.25 |
| N°2 | 79.44 | 71.96 |
| N°3 | 80.56 | 72.22 |
| N°4 | 61.02 | 38.14 |
| N°5 | 50.00 | 23.44 |
| Average | 70.45 | 56.40 |
Table 7: Egg incubation success per nest.
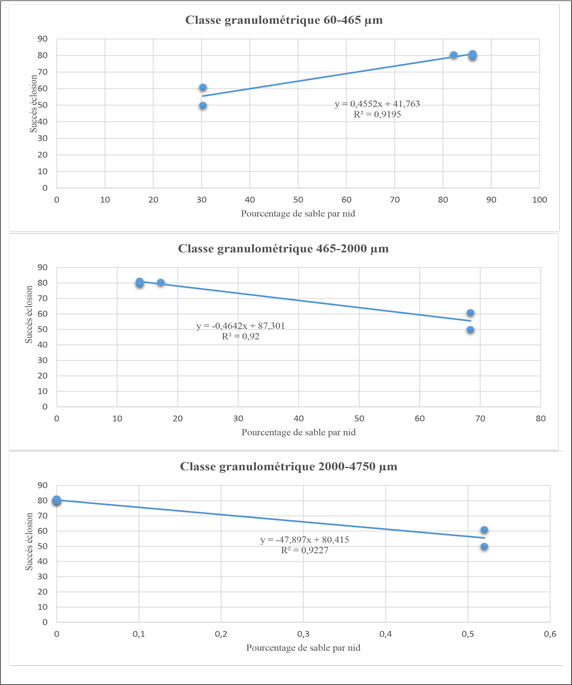
Correlation between incubation success and particle size
The analysis of Table 7 reveals in the particle size range 60-465µm (fine sand) a positive and strong correlation with hatching success (r²=0.95) and emergence success (r²=0.97). On the other hand, in the particle size ranges 465-2000 µm (medium sand) and 2000-4750 µm (coarse sand) the analysis reveals a negative and strong correlation with hatching success and emergence success, which allows us to say that there is a significant link between particle size and incubation success.
The analysis of Table 7 reveals in the particle size range 60-465µm (fine sand) a positive and strong correlation with hatching success (r²=0.95) and emergence success (r²=0.97). On the other hand, in the particle size ranges 465-2000 µm (medium sand) and 2000-4750 µm (coarse sand) the analysis reveals a negative and strong correlation with hatching success and emergence success, which allows us to say that there is a significant link between particle size and incubation success.
| Incubation | Granulometry | ||
| 60-465 | 465-2000 | 2000-4750 | |
| Hatching | 0.95** | - 0.95** | - 0.96** |
| Emergence | 0.97** | - 0.97** | - 0.97** |
Table 8: Correlations of particle size with incubation success.
Linear regression of incubation success as a function of particle size
The correlation test showed that there is a link between particle size and incubation success, which allows us to identify a linear regression of incubation success as a function of particle size. Figure 7 shows the linear regressions of hatching and emergence success as a function of the percentage of sand of the particle size classes.
The correlation test showed that there is a link between particle size and incubation success, which allows us to identify a linear regression of incubation success as a function of particle size. Figure 7 shows the linear regressions of hatching and emergence success as a function of the percentage of sand of the particle size classes.
Figure 7: Linear regression of hatching success as a function of the percentage of sand by particle size class.
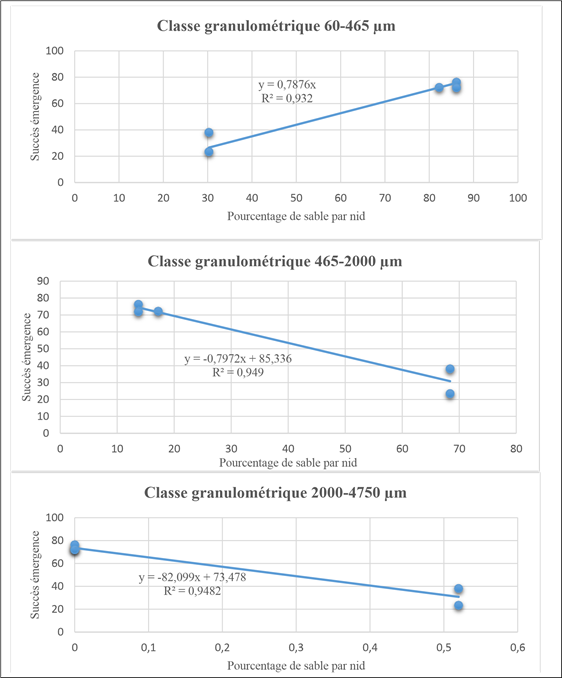
Figure 8 below shows the linear estimation of emergence success by particle size class. Each class constitutes a category of sand. Overall, it appears that emergence success is low when the size of the sediment increases.
Figure 8: Linear regression of emergence success as a function of percentage of sand particle size class.
Discussion
Composition of woody species and animal species encountered on olive ridley turtle nesting sites
The Cameroonian coast is particularly rich in numerous faunal and floral species and particularly marine turtles which frequent this coastline for feeding and nesting Angoni, 2005, Ayissi, 2000 et Angoni et al, 2010. Thus, the results on the animal species of this study show that the nesting sites are composed of several animal species, namely crabs, mammals (dogs, civet, pangolin and monkeys), birds and some reptiles such as the monitor lizard and the Agama lizard. These species have been observed at nesting sites because they are predators of eggs and hatchlings. The results are similar, for example, to those described in French Guyana, on the Yalimapo nesting beach and the beaches of Basse-Mana for predation on turtle eggs and hatchlings Fretey, 1977, Fretey, 2005 et Fretey & Frenay, 1980. These report that amerindians stray dogs, crab raccoons, birds (nocturnal and diurnal), crabs and mole crickets frequent the coastal area, the nesting site of sea turtles and are predators of the emerging hatchlings on the beaches. Like this author, Maite et al. 2008 in his studies on the nesting of the leatherback turtle in the Pongara National Park in Gabon mentions the presence of species such as: Nile monitor lizards, mongooses, insects (ants, termites, beetles), crabs and others predators of sea turtles.
The Cameroonian coast is particularly rich in numerous faunal and floral species and particularly marine turtles which frequent this coastline for feeding and nesting Angoni, 2005, Ayissi, 2000 et Angoni et al, 2010. Thus, the results on the animal species of this study show that the nesting sites are composed of several animal species, namely crabs, mammals (dogs, civet, pangolin and monkeys), birds and some reptiles such as the monitor lizard and the Agama lizard. These species have been observed at nesting sites because they are predators of eggs and hatchlings. The results are similar, for example, to those described in French Guyana, on the Yalimapo nesting beach and the beaches of Basse-Mana for predation on turtle eggs and hatchlings Fretey, 1977, Fretey, 2005 et Fretey & Frenay, 1980. These report that amerindians stray dogs, crab raccoons, birds (nocturnal and diurnal), crabs and mole crickets frequent the coastal area, the nesting site of sea turtles and are predators of the emerging hatchlings on the beaches. Like this author, Maite et al. 2008 in his studies on the nesting of the leatherback turtle in the Pongara National Park in Gabon mentions the presence of species such as: Nile monitor lizards, mongooses, insects (ants, termites, beetles), crabs and others predators of sea turtles.
According to our study, the tree composition of nesting sites shows that they are composed of several woody species (09) divided into families (06), namely: Calophylllum inophyllum, Ficus microcarpa, Cocos nucifera, Terminalia catapa, Musanga cecropioides, Ficus saussureana, Phoenix Spp, F. benjamina and Elaeis guineensis. These results seem to be similar to those found by Angoni, 2005 on the vegetation of the seafront of the Kribi-Campo coastal strip. Indeed, he reports that the vegetation of the Kribi-Campo seafront is populated by species such as Manilkara obovata, Calophyllum inophyllum, Terminalia catapa and Pentadesma butyracea. Further back we find: Barteria nigritiana, Cuviera longifolia, Anthonotha sp., Oddoniodendron sp. This slight difference in results may be due to the geographical position and the number of study plots and also the nesting sites studied.
The most abundant woody species in this study is Calophyllum inophyllum, with an average density equal to 58.35%, as it is present on all sites. This result differs from that obtained by Dossa et al. 2007 in his study of marine turtle populations and their nesting habitat on the coast of Grand-Popo in Benin. Indeed, the most abundant species obtained in this work is Cocos nucifera with an average density equal to 36%, compared to an average density of 9.13% in the present study, this difference in results can be explained by the methodology used. Indeed, transects used in these studies cross plant formations including fallow, degraded thicket, degraded mangrove, coconut grove and coastal grassland.
Nature of the substrate and physical characteristics nesting sites.
The substrate identified in this study is homogeneous and sandy in nature and the grain size analysis reveals fine to coarse sediments. These results obtained are similar to those found by Angoni, 2005 in his study on the biology and ecology of marine turtles in relation to coastal ecosystems in the Kribi-Campo area. However, these two results are very different from those found by Peron, 2014 in his study of coastal dynamics and sea turtle behaviour in French Guiana on the Awala-Yalimapo nesting beach. He reports that the spawning substrate is sand associated with clay particles.
The substrate identified in this study is homogeneous and sandy in nature and the grain size analysis reveals fine to coarse sediments. These results obtained are similar to those found by Angoni, 2005 in his study on the biology and ecology of marine turtles in relation to coastal ecosystems in the Kribi-Campo area. However, these two results are very different from those found by Peron, 2014 in his study of coastal dynamics and sea turtle behaviour in French Guiana on the Awala-Yalimapo nesting beach. He reports that the spawning substrate is sand associated with clay particles.
The Pearson correlation test showed the link of particle size with incubation success, there is a strong correlation of particle size with incubation success (r²=0.95 for hatching and r²=0.97 for emergence) moreover, incubation success is high when the sand is predominantly fine, that is to say in the particle size range 60-465 µm. The results obtained by Peron, 2014 on the nesting beach of Awala-Yalimapo in French Guiana show that there is a strong correlation of the particle size gradient for the situation of leatherback turtle and green turtle nests (r²=0.98, p < 0.05), and the preferential particle size range for laying eggs for these two species goes from 355 to 200 μm.
Conclusion
The results of this study carried out in the Manyange na Elombo-Campo National Marine Park on certain biological and physical characteristics reveal that marine turtle nesting sites are made up of several well-defined woody species. Eggs and new born marine turtles are predated by various invertebrate and vertebrate animal species such as crabs, mammals, birds and reptiles.
A total of nine woody species divided into six different families, namely the Arecaceae, Clusiaceae, Combretaceae, Moraceae, Sapotaceae and Urticaceae were observed in these egg-laying sites. The analysis of the substrate of the nesting sites shows that they are made up of a homogeneous substrate of a sandy nature but with the topo-morphology which varies depending on the nesting sites. The grain size was more than 60% made up of fine sand in all the sites studied. Incubation success was inversely proportional with increasing sand seed size.
Acknowledgements
All authors thanks to the local Community Research and Development Association “TUBE AWU” in Ebodje, in southern Cameroon for their help in data collection.
All authors thanks to the local Community Research and Development Association “TUBE AWU” in Ebodje, in southern Cameroon for their help in data collection.
References
- Angoni, H., (2005). Biologie et Écologie des tortues Marines en rapport avec les écosystèmes côtiers. Conservation et Aménagement. Thèse de Doctorat 3è Cycle, Université de Yaoundé I, 125 p.
- Angoni, H., Amougou, A., Bilong, B. C. F. & J. Fretey, J., (2010). La tortue marine au Cameroun, genre Lepidochelys: nidification, biométrie de Lepidochelys olivacea (Eschscholtz, 1829) (Reptilia, Cheloniidae) dans la Réserve de Faune de Campo-Ma’an (Sud Cameroun). Int. J ; Biol. Chem. Sci., 4 (3) : 649-656.
- Ayissi I., (2000). Projet d’un Ecotourisme «Tortues marines» dans la Région d’Ebodjé. (Unité Technique Opérationnelle de Campo-Ma’an). Mémoire DESS, Université de Yaoundé I, 57 p.
- Bell, B. A., Spotila, J. R., Paladino, R. V. & Reina, R. D., (2003). Low reproductive success of Leatherback turtle, Dermochelys coriacea, is due to embryonic mortality. Biological Conservation, 131-138.
- Bjorndal, K. A. & J. B. C. Jackson, (2003). Roles of sea turtles in marine ecosystems: reconstructing the past. Pp. 259-273 In: Lutz, P L, Musick, J. & J. Wyneken (Eds.), Biology of Sea Turtles, volume II. CRC Press, Boca Raton, FL, 472 p.
- Bjorndal, K. A., Bolten A. B, Chaloupka, M. Y., (2000). Modèle de croissance somatique de la tortue verte : preuve de la dépendance de la densité. Ecol Appl., 10: 269-282.
- Bourgea, J., (2014). Structure et connectivité de la mégafaune marine à l’échelle d’une région océanique : enjeux pour la gestion durable des tortues vertes dans l’océan Indien occidental. Thèse de doctorat en Sciences de la Vie-Ecologie, Université de la Réunion, 169 p.
- Defeo O., McLachlan, A., Schoeman, D., Schlacher, T. A., Dugan, J., Jones, A., Lastra, M. & F. Scapini, (2009). Menaces sur les écosystèmes des plages de sable: une étude. Science des côtes et du plateau continental estuarien, 81 :1-12.
- Dessouassi, C. E., Lederoun, D., Chikou, A. & P.A. Laleye., (2018). Diversité et caractéristiques des crabes de lagunes au Sud-Bénin, Afrique de l'Ouest. Afrique Science, 14(6) : 366-387.
- Dossa, S. J., Mensah, G A. & A. Laleye, (2007). Etude des populations de tortues marines et leur habitat de ponte sur la côte de Grand-Popo. Bulletin de la Recherche Agronomique du Bénin. 55 : 17-24.
- Fretey, J. & P. Triplet, (2021). Ramsar sites and marine turtles an overview. Ministère de la Transition écologique & Chélonée, 244 p.
- Fretey, J. (Coord.), (2005). Les tortues marines de Guyane. Ed. Plume Verte, 190 p.
- Fretey, J., & D. Frenay, (1980). Prédation des nids de Tortues luth (Dermochelys coriacea) par les chiens des villages indiens Galibi en Guyane française. Rev. Méd. vétér., 131(12): 861-867.
- Fretey, J., & M. Girondot, (1996). Mise au point d'une fiche de description de sites de ponte. Rapport Ministère de l'Environnement, Direction de la nature et des paysages. 15 p.
- Fretey, J., 1977. Causes de mortalité des Tortues luths adultes (Dermochelys coriacea) sur le littoral guyanais. Courr. Nature, 52 : 257-266.
- Fretey, J., (2003). Lexique zoologique français /iyassa-Les Vertébrés. Mimeogr. UICN, Projet Campo Ma’an, 20 p.
- Fuentes, M. M. P. B., Mariana M. P. B., McMichael, E., Kot, C. Y., Silver-Gorges, I., Wallace, B. P., Godley, B. J., Brooks, A. M. L., Ceriani, S. A., Cortés-Gómez, A. A., Dawson, T. M., Dodge, K. L., Flint, M., Jensen, M. P., Komoroske, L.M., Kophamel, S., Lettrich, M.D., Long, C. A., Nelms, S. E., Patrício, A. R., Robinson, N. J., Seminoff, J. A., Ware, M., Whitman, E. R., Chevallier, D., Clyde-Brockway, C. E., Korgaonkar, S. A., Mancini, A., Mello-Fonseca, J., Monsinjon, J. R., Neves-Ferreira, I., Ortega, A. A., Patel, S. H., Pfaller, J. B., Ramirez, M. D., Raposo, C., Smith, C. E., Abreu-Grobois, F. A. & C. G. Hays, (2023). Key issues in assessing threats to sea turtles: knowledge gaps and future directions. Endangered Species Research, 52: 303-341.
- Maite, I. S. & D.P. Agambouet, (2008). Trois Années d´Étude sur la nidification de la Tortue Luth au Parc National de Pongara, Gabon, Afrique Centrale-Synthèse d’activités, Avril 2008. Rapport, 42 p.
- Manning, R.B. & Holthuis, L.B., (1981). West African brachyuran crabs (Crustacea: Decapoda). Smithsonian Contributions to Zoology. 306: 1-379., available online at http://www.sil.si.edu/smithsoniancontributions/zoology/pdf_hi/SCTZ-0306.pdf
- Peron C., 2014. Dynamique littorale et comportement de ponte des tortues marines en Guyane française. Géomorphologie. Université du Littoral Côte d’Opale, 226 p.
- UICN, (2012). Cartographie des sites de ponte des tortues marines en Afrique de l’ouest, pp. ????
- Wood, D. & K. A. Bjorndal, (2000). Relation of Temperature, Moisture, Salinity, and Slope to Nest Site Selection in Loggerhead Sea Turtles. Copeia. 1: 119-128.
Citation: Nana Towa Algrient, Mondjeli Ndjokou Djongo Lampert Yves Maxime, Ayissi Isidore and Fretey Jacques. (2025). Biophysical characterization of nesting sites of olive ridley turtle (Lepidochelys olivacea, Eschsholtz, 1829) in the «Manyange na Elombo-Campo» National Marine Park, South Cameroon (West-Africa). Journal of Agriculture and Aquaculture 7(1).
Copyright: © 2025 Nana Towa Algrient. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.